Mutation is a change in the DNA sequence of an organism’s genetic material. DNA, or deoxyribonucleic acid, is the molecule that carries the genetic instructions for the development and function of all living things.
DNA is made up of four chemical bases, adenine (A), cytosine (C), guanine (G), and thymine (T), which are arranged in a specific sequence to form genes. Genes are the functional units of DNA that encode specific traits and are passed on from parent to offspring.
In biology, mutations can occur naturally or as a result of exposure to certain environmental factors such as radiation or chemicals. Natural mutations can arise from errors during DNA replication or due to spontaneous chemical changes in DNA. Mutations can also be introduced into the DNA through infection by viruses, which can insert their own genetic material into the host cell.
Mutations can have a variety of effects on the organism. Some mutations may have no effect and are referred to as silent mutations. These mutations are often located in non-coding regions of the DNA and do not affect the function of the gene. Other mutations may result in a change in the amino acid sequence of a protein, which can cause it to lose function, gain new function, or alter its activity.
Some mutations can be harmful and cause diseases such as cancer, genetic disorders, and developmental disorders. For example, mutations in the genes that regulate cell division can lead to uncontrolled cell growth and the formation of a tumor. Mutations in genes involved in the development of the nervous system can result in developmental disorders such as autism and intellectual disability.
However, mutations can also have beneficial effects and play a role in evolution. Beneficial mutations are those that increase an organism’s fitness and enable it to better adapt to its environment. Over time, these mutations can accumulate and lead to the development of new species. This process, known as natural selection, is the driving force behind evolution.
Mutations can occur in different parts of the DNA sequence. Point mutations, for example, involve the substitution of one base for another. Frameshift mutations are caused by the insertion or deletion of one or more bases in the DNA sequence, which changes the reading frame of the gene and can result in a complete change in the amino acid sequence of the encoded protein.
Mutations can also occur in regulatory regions of the DNA, such as enhancers and promoter regions, which control the expression of a gene. These mutations can result in changes in gene expression levels, leading to alterations in the pattern of gene expression and cellular behavior.
In summary, mutations play a crucial role in biology, both as a source of genetic variation and as a driving force behind evolution. They can result in both positive and negative effects on an organism, and the impact of a mutation depends on the specific location, type, and frequency of the mutation. The study of mutations and their effects on organisms is a central focus of genetics and evolutionary biology and is important for understanding the development of diseases and the mechanisms of evolution.

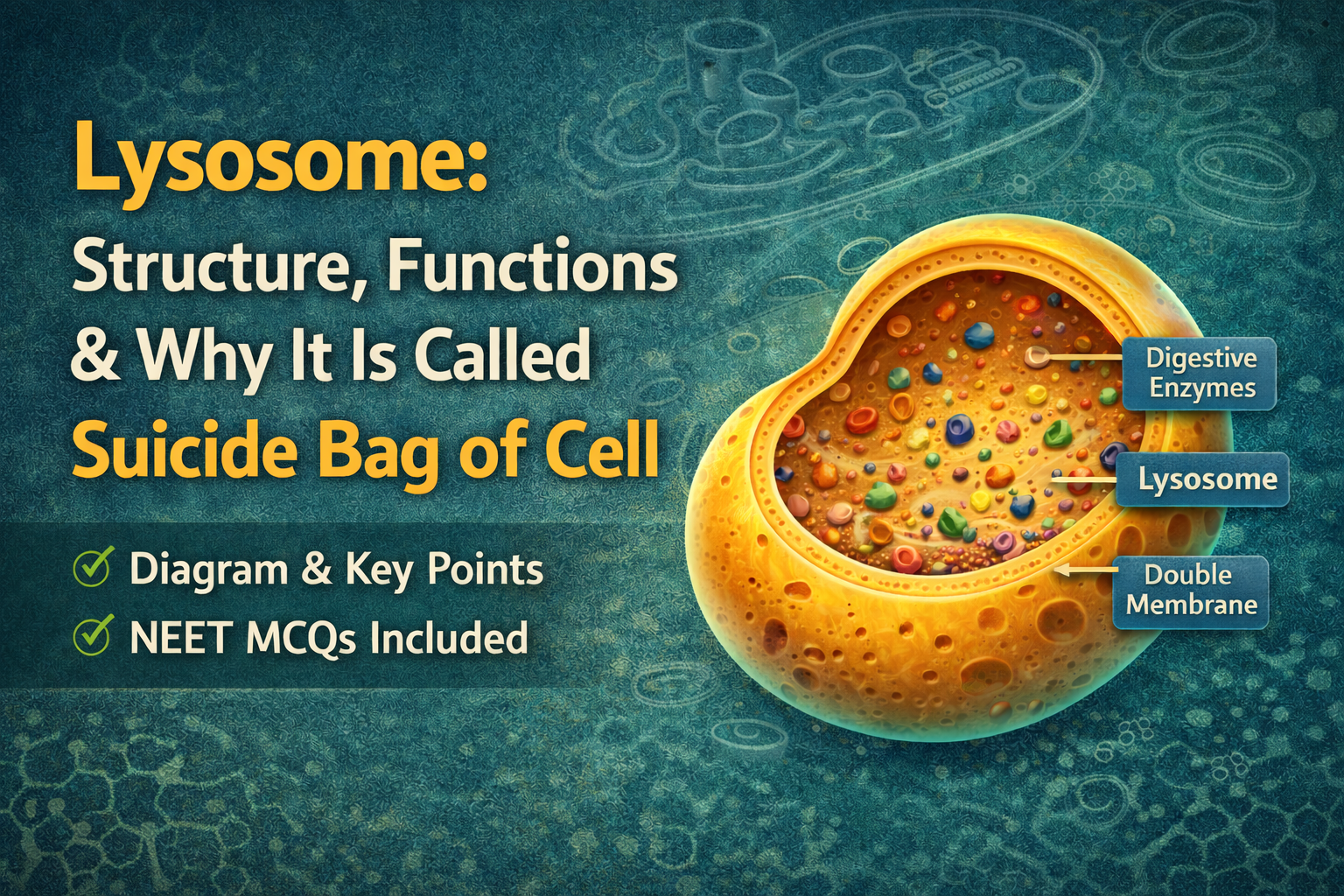
[…] Mutation […]