
Scientific name – Balanoglossus
Common name – Acorn worm or Tongue worm
Class – Enteropneusta
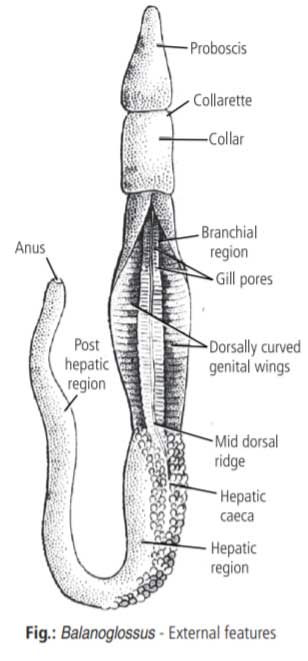
Balanoglossus clavigerus was recorded and named by Delle Chiaje in 1829.
Its generic name was derived from two Greek words, balanos and glossus. Term balanos means an “acorn” (fruit of oak) and refers to the proboscis projecting from collar looking like an acorn-nut, hence the common name “acorn worm”. Term glossus means “tongue” and refers to the shape of its proboscis, collar and genital wings bearing a close resemblance to an ox tongue, hence the common name “tongue worm”.
Balanoglossus is a marine, tubicolous or burrowing hemichordate inhabiting shallow coastal waters of intertidal zone.
Body is soft, elongated, worm-like, cylindrical and bilaterally symmetrical but unsegmented. It is divided into anterior-most proboscis or protosome, middle collar and posterior long trunk with terminal anus.
Proboscis appears as a tongue-like projection.
Collar is the muscular region which bears mouth ventrally below the proboscis stalk.
Trunk is divisible into three regions :
- The anterior branchiogenital region which has gill pores and gonads. The lateral regions containing the gonads are thin and flat and form genital wings.
- The middle hepatic region is marked externally with irregular elevations due to sacculations formed by projecting hepatic caeca of the intestine.
- The posterior abdominal region gradually tapers posteriorly and bears a terminal anus.
Coelom is spacious and lined by coelomic epithelium and is enterocoelous in origin.
The animal lives inside a U-shaped tube or burrow with two vertical limbs and two openings.
The worm moves in its burrow by cilia covering its body surface although the worm is sluggish and little affected by external stimuli. The proboscis is the most active part which helps in burrowing.
Balanoglossus feeds on diatoms, protozoans, other microorganisms and organic detritus by swallowing sand or mud.
Balanoglossus is a ciliary feeder and the lateral cilia lining the gill slits set up a current of water which enters through the mouth, takes its course through the buccal cavity, pharynx, gill slits and branchial sacs and leaves through the gill pores. This is known as respiratory-cum-food current.
Respiratory apparatus of Balanoglossus comprises : (i) branchial portion of pharynx bearing gill-slits, and (ii) branchial sacs that open out through gillpores.
Blood vascular system of Balanoglossus is of the open or lacunar type. It consists of : (i) a colourless blood, (ii) a central sinus and a heart vesicle, (iii) distributing vessels or arteries and sinuses, and (iv) collecting vessels or veins.
The excretory organ is glomerulus or proboscis gland lying in front of the central sinus and projecting into the proboscis coelom. It excretes brown or yellow granules.
Nervous system is of primitive type resembling that of coelenterates and echinoderms.
Sexes are separate i.e., dioecious and males and females live in separate tubes. Fertilisation is external as gametes are shed in sea water and fuse there.
Development is indirect with a free swimming planktonic ciliated larva, the tornaria larva.
It does not reproduce asexually but it has great power of regeneration.
Scientific name – Saccoglossus
Common name – Tongue worm
Class – Enteropneusta
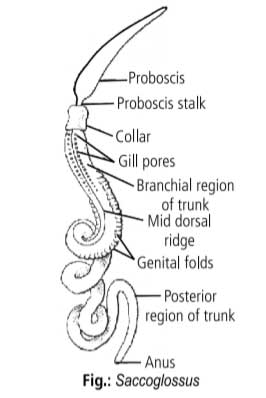 It is a typical enteropneust genus very much similar to Balanoglossus in habitat, habits and structure.
It is a typical enteropneust genus very much similar to Balanoglossus in habitat, habits and structure.
It is a marine, slender, soft-bodied tubicolous tongue worm living in spirally twisted burrows. Body has the usual three divisions – proboscis, collar and trunk.
Genital wings and hepatic caeca are absent. Mature gonads are yellow in male and grey in female and their position marked externally by dorsolateral genital folds in the middle part of the trunk.
Development is direct without a free-swimming tornaria larva.
Saccoglossus pygmaeus, measuring 2 to 3 cm in length, represents the smallest known species of Enteropneusta.

Leave a Reply
You must be logged in to post a comment.