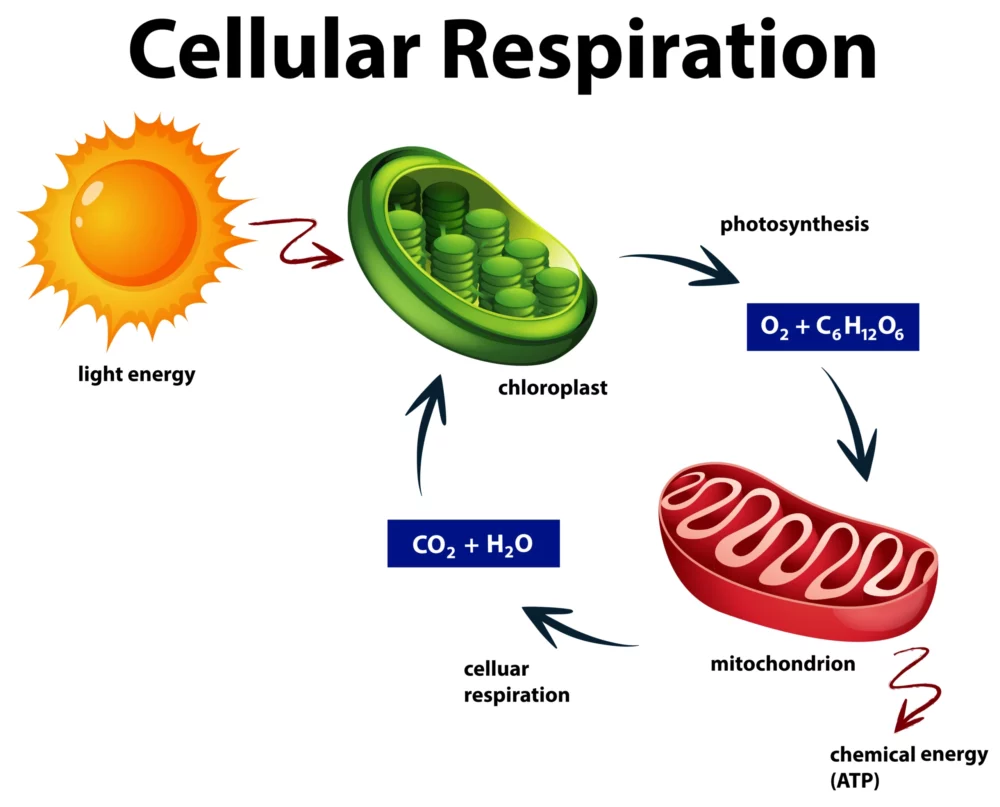
Cellular respiration is a fundamental biochemical process that occurs in all living cells, from the tiniest bacteria to the most complex multicellular organisms. It is the means by which cells convert organic molecules, primarily glucose, into adenosine triphosphate (ATP) – the universal currency of cellular energy. This intricate and highly regulated metabolic pathway is essential for life, providing the energy necessary for various cellular functions and processes.
Stages of Cellular Respiration
Cellular respiration occurs in multiple stages, each with specific enzymes and reactions involved:
Glycolysis:
Glycolysis is the first stage of cellular respiration and takes place in the cytoplasm. It involves the breakdown of one molecule of glucose (a six-carbon compound) into two molecules of pyruvate (a three-carbon compound). This process does not require oxygen and yields a small amount of ATP and NADH (a coenzyme carrying electrons) as energy carriers.
Pyruvate Decarboxylation:
In the presence of oxygen, pyruvate produced during glycolysis enters the mitochondria. There, it undergoes a series of reactions, including decarboxylation, to form Acetyl-CoA, releasing carbon dioxide in the process. This step connects glycolysis to the next stages of respiration.
Citric Acid Cycle (Krebs Cycle):
The citric acid cycle takes place in the mitochondria’s matrix. Acetyl-CoA enters the cycle, where it is oxidized, releasing high-energy electrons and carbon dioxide. This cycle generates ATP, NADH, and FADH2 (another electron carrier), and it serves as a central hub in cellular respiration.
Electron Transport Chain (ETC):
The electron transport chain is located in the inner mitochondrial membrane. It is where the high-energy electrons carried by NADH and FADH2 are transported through a series of protein complexes. As electrons move through the chain, energy is released, which is used to pump protons (H+) across the inner mitochondrial membrane, creating an electrochemical gradient.
Chemiosmotic ATP Synthesis:
The proton gradient created by the ETC drives protons back into the mitochondrial matrix through ATP synthase complexes, a process known as chemiosmosis. This flow of protons powers the synthesis of ATP from adenosine diphosphate (ADP) and inorganic phosphate (Pi), a process called oxidative phosphorylation.
The Role of Oxygen
Oxygen plays a crucial role in cellular respiration. It serves as the terminal electron acceptor in the electron transport chain. Without oxygen, the ETC and chemiosmotic ATP synthesis cannot occur efficiently, leading to a reduced yield of ATP and a shift towards less efficient forms of energy production.
Energy Yield
Cellular respiration is highly efficient at extracting energy from glucose. The complete oxidation of one molecule of glucose through cellular respiration can yield up to 36-38 molecules of ATP, depending on various factors. This energy is essential for powering various cellular processes, including muscle contraction, active transport, and biosynthesis of molecules.
Anaerobic Respiration
In the absence of oxygen, cells can resort to anaerobic respiration, which includes processes like lactic acid fermentation or alcoholic fermentation. While these pathways can generate ATP, they are less efficient and can lead to the accumulation of metabolic byproducts like lactic acid or ethanol.
Conclusion
Cellular respiration is the biochemical process that fuels life. By converting glucose and other organic molecules into ATP, cells obtain the energy needed to perform their functions and maintain life processes. This intricate series of metabolic reactions, from glycolysis to the electron transport chain, exemplifies the elegance and complexity of biological systems. Understanding cellular respiration is not only vital for biology but also for appreciating the marvel of life’s energy dynamics.

Leave a Reply
You must be logged in to post a comment.